Fire Ecology I
![]() No.
54, November/December 2003
No.
54, November/December 2003
Fire Ecology I
by María del Carmen COBO and José Antonio CARREIRA
| "Our study provides solid results to interpret the most important processes involved in the mid- to long-term evolution of litter and soil nutrient contents and pools in Mediterranean shrubland ecosystems in relation to changes in attributes of the fire perturbation regime (frequency and time elapsed since the last fire), which underlie desertification processes in fire-prone, semiarid areas." |
Introduction(Back to top) Conceptual framework |
|
|
(Back to top) We hypothesize, however, that when fire frequency is abnormally high, the ecosystem nutrient capital does not fully recover, and the long-term balance between fire-related nutrient outputs and succession-related nutrient inputs is upset. Long-lasting soil fertility depletion slows rates of vegetation recovery which, in turn, increases erosion rates, initiating a positive feedback loop that can lead to irreversible desertification (Boerner 1982; Schlesinger et al. 1990; Carreira, Niell and Lajtha 1994). Our objective(Back to top) The experimental system |
|
|
(Back to top) |
|
|
The study sites consisted of a four-stand, soil and vegetation chronosequence (Dyck and Cole 1990) of recent fire history. The sampling plots were selected within each of these study sites (four adjacent, steep sub-basins 1 to 3 km apart), all in mid-hillside positions with similar elevation (300-400 m), exposure (SE) and slope (36-41º). Prior to an initial "baseline" fire (1975), the whole study area was covered by a uniform old-growth Pinus pinaster forest with dense garrigue understory. |
|
|
After 1975, the four study sites chosen, each 1000 m2, differed in their fire-disturbance regime (fire-frequency and time-lag between fire events): Unit D had not burned during the 1975 baseline fire or indeed since at least the 1950s, and was used as the reference plot. The three other plots, Units A, B, and C, burned in 1975; of these, Units A and B burned a second time in 1981, and Unit A burned a third time in 1986 (that is, Unit D had 0 fires, Unit C had 1 fire, Unit B had 2 fires, and Unit A had 3 fires during the period 1975 to 2002). Current species composition is similar in all units, with different proportions of abundance. Vegetation on relatively well-developed soils consists of discontinuous stands of garrigue shrublands with Juniperus oxycedrus, Buxus balearicae and Pistacia lentiscus as dominant species, and open pine (Pinus pinaster) forests. Where frequent fires have occurred, garrigue formations are replaced by gorse (Ulex rivasgodayanus) and rockrose (Cistus clusii)-dominated dwarf-shrub vegetation that includes many dolomitophilous plant endemics (Carreira, Lajtha and Niell 1997). Soil collection and analyses |
|
|
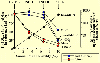
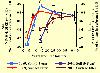
(Back to top) Within 48 h after collection, litter and soil sub-samples were taken apart to calculate field moisture. Next, the litter samples were cleaned and air-dried. The soil samples were air-dried and sieved to 2 mm. Litter mass, soil gravel content and bulk density were recorded. After that, general soil chemical properties and total and labile nutrient (C, N and P) contents were measured. For both sampling periods, litter and soil nutrient contents and pool sizes were expressed as a percentage of their value at the reference sampling plot (Unit D), and represented as a function of the number of years passed since the last fire event. During the 1989-1991 sampling period, we also measured soil erosion rates using Verlagh boxes (Carreira, 1992). Nutrient pool sizes in litter and soil from this period were expressed as a percentage of their value at the reference plot (Unit D), together with erosion fluxes expressed as number of times above values at the reference plot, and represented as a function of site annual fire probability over the last 45 years. What was found? |
|
|
(Back to top) |
|
|
Another interesting issue that arose from our data relates to an alternative meaning for the concept of stability: elasticity (Grimm and Wissel 1997; Brang 2001). The soil nutrient pool at Unit A, the sampling plot that experienced an abnormally short fire return interval (3 fires) during 1975-1986, showed very small or even no recovery during the interval from 1989/91 (3-5 yr-old vegetation at Unit A, surface soil total N pool size about 30% of that in the reference plot) to 2000/02 (14-16 yr-old vegetation at Unit A, soil N pool size 35% of that in the reference plot), even though no new fires had taken place. In contrast, the surface soil total N pool size at Unit C (burned once), when its vegetation was 14-16 yr-old (in the 1989-91 period), was 125% of that in the reference plot. Thus, in the long term, the size of the soil nutrient capital depends more on the fact that a given threshold of fire recurrence has been surpassed at some time in the past, than on the time elapsed since the last fire event took place. This means that the soil nutrient pool, despite its relatively high inertia against fire, shows very low elasticity (that is, there are long-lasting effects if fire recurrence is high). On the other hand, the litter N content in Unit A, that was just 45% of N content in the reference plot during the first sampling period (1989-91), increased to 70% of reference values during the period 2000-02. This outstanding recovery for the litter N contents in the absence of new disturbance events, in comparison to the slight increments observed for the soil N contents and pools in the same circumstances, demonstrates the higher elasticity, although lower inertia, of nutrients in the litter layer. Conclusions(Back to top) We found that erosion fluxes showed an exponential-type response curve with respect to a linear-increasing annual fire probability at the study area. However, when nutrient pools were considered, a threshold-type response curve was found. Above a given fire-frequency threshold, differences among sampling plots were more strongly related to the number of past fires (fire frequency) than to the age of the vegetation stand (number of years since the last fire). Our study indicated that in sampling plots experiencing an abnormally short fire-return interval, even transitory, during the last five decades, the current post-fire recovery rate of the nutrient capital and its associated process-rates was very low or is even absent (sensu positive degradation feedback leading to desertification; Schlesinger et al. 1990). On the other hand, in the sampling plots not surpassing such a threshold, recovery of the nutrient capital or organic matter cycling process-rates was always achieved in the short to mid-term, or is even increased with respect to the reference plot. This, in turn, stresses that perturbation is necessary for ecosystem sustainability, provided that a given perturbation regime is maintained through time and perturbation thresholds are not surpassed (Holling 1986; Chapin, Torn and Tatano 1996). References(Back to top) Boerner, R.E.J. 1982. Fire and nutrient cycling in temperate ecosystems. Bioscience 32: 187-192. Brang, P. 2001. Resistance and elasticity: Promising concepts for the management of protection forests in the European Alps. Forest Ecology and Management 145: 107-119. Carreira, J.A. 1992. Biogeoquímica y disponibilidad del N y P en una cronosecuencia de suelos semiáridos dolomíticos asociada a distintos regímenes de incendios. Doctoral dissertation, University of Málaga, Spain. Carreira J.A., F.X. Niell and K. Lajtha. 1994. Soil nitrogen availability and nitrification in Mediterranean shrublands of varying fire history and successional stage. Biogeochemistry 26: 189-209. Carreira, J.A., J.R. Arévalo and F.X. Niell. 1996. Soil degradation and nutrient availability in fire-prone Mediterranean shrublands of southeastern Spain. Arid Soil Research and Rehabilitation 10: 53-64. Carreira, J.A., K. Lajtha and F.X. Niell. 1997. Phosphorus transformations along a soil/vegetation series of fire-prone, dolomitic, semi-arid shrublands of southern Spain. Biogeochemistry 39: 87-120. Chapin, F.S., M.S. Torn and M. Tateno. 1996. Principles of ecosystem sustainability. The American Naturalist 148: 1016-1037. Christensen, N.L. 1977. Fire and soil-plant nutrient relations in a pine wiregrass savanna on the Coastal Plain of North Carolina. Oecologia 31: 27-44. DeBano, L.F. and C.E. Conrad. 1978. The effect of fire on nutrients in a chaparral ecosystem. Ecology 59: 489-497. De Groot, W.J., P.M. Bothwell, D.H. Carlsson and K.A. Logan. 2003. Simulating the effects of future fire regimes on western Canadian boreal forests. Journal of Vegetation Science 14 (3): 355-364. Dunn, P.H., L.F. DeBano and G.E. Eberlein. 1979. Effects of burning on chaparral soils: II. Soil microbes and nitrogen mineralization. Soil Science Society of America Journal 45: 504-514. Dyck, W.J. and D.W. Cole. 1990. Requirements for site productivity research. In Impacts of intensive harvesting on forest site productivity, ed. W.J. Dyck and C.A. Mees. Proceedings of the IEA/BE A3 Workshop, Marlborough Sounds, New Zealand, March 1989. IEA/BE T6/A6 Report No. 2. Rotorua, New Zealand: Ministry of Forests, Forest Research Institute. Fuhlendorf, S., F. Smeins and W. Grant. 1996. Simulation of a fire-sensitive ecological threshold: A case study of Ashe juniper on the Edwards Plateau of Texas, USA. Ecological Modelling 90: 245-255. Grimm, V. and C. Wissel. 1997. Babel, or the ecological stability discussions: An inventory and analysis of terminology and a guide for avoiding confusion. Oecologia 109: 323-334. Hanes, T.L. 1971. Succession after fire in the chaparral of southern California. Ecological Monographs 41: 27-52. Holling, C.S. 1973. Resilience and stability in ecological systems. Annual Review of Ecology and Systematics 4: 1-23. Holling, C.S. 1986. The resilience of terrestrial ecosystems: Local surprise and global change. In Sustainable development of the biosphere, ed. W. Clark and R. Mund, 292-317. Cambridge, UK: Cambridge University Press. Johnson, D.W., R.B. Susfalk, R.A. Dahlgren and J.M. Klopatek. 1998. Fire is more important than water for nitrogen fluxes in semi-arid forests. Environmental Science and Policy 1: 79-86. Meyer, G.A. and J.L. Pierce. 2003. Climatic controls on fire-induced sediment pulses in Yellowstone National Park and central Idaho: A long-term perspective. Forest Ecology and Management 178 (1-2): 89-104. Muradian, R. 2001. Ecological thresholds: A survey. Ecological Economics 38: 7-24. Naveh, Z. 1989. Fire in the Mediterranean: A landscape ecological perspective. In Fire in Ecosystem Dynamics, ed. J.G. Goldammer and J. Jenkins, 1-20. Proceedings of the Third International Symposium on Fire Ecology, Freiburg, FRG, May 1989. Neary, D.G., C.C. Klopatek, L.F. DeBano and P.F. Ffolliott. 1999. Fire effects on belowground sustainability: A review and synthesis. Forest Ecology and Management 122 (1-2): 51-71. Nieto, J.M. and B. Cabezudo. 1988. Series de vegetación climatófilas de las Sierras de Tejeda y Almijara (Málaga-Granada, España). Acta Botanica Malacitana 13: 229-260. Nieto, J.M., Cabezudo B. and M.M. Trigo. 1989. Series de vegetación edafófilas de las Sierras de Tejeda y Almijara (Málaga-Granada, España). Acta Botanica Malacitana 14: 161-170. Orians, G.H. 1975. Diversity, stability and maturity in natural ecosystems. In Unifying concepts in ecology, ed. W.H. van Dobben and R.H. Lowe-McConnell, 139-150. The Hague: Dr. W. Junk BV Publishers. Romanyà, J., P. Casals and V.R. Vallejo. 2001. Short-term effects of fire on soil nitrogen availability in Mediterranean grasslands and shrublands growing in old fields. Forest Ecology and Management 147: 39-53. Rundel, P.W. and D.J. Parsons. 1980. Nutrient changes in two chaparral shrubs along a fire-induced age gradient. American Journal of Botany 67: 51-58. Rundel, P.W. 1983. Impact of fire on nutrient cycles in Mediterranean-type ecosystems with reference to chaparral. In Mediterranean-type ecosystems: The role of nutrients, ed. F.J. Kruger, D.T. Mitchel and J.V.M. Jarvis, 192-207. Berlin: Springer-Verlag. Scarascia-Mugnozza, G., H. Oswald, P. Piussi and K. Radoglou. 2000. Forests of the Mediterranean region: Gaps in knowledge and research needs. Forest Ecology and Management 132: 97-109. St. John, T.V. and P. W. Rundel. 1976. The role of fire as a mineralizing agent in a Sierran coniferous forest. Oecologia (Berl.) 25: 35-45. Schlesinger, W.H., J.F. Reynolds, G.L. Cunningham, L.F. Huenneke, W.M. Jarrel, R.A. Virginia and W.G. Whitford. 1990. Biological feedbacks in global desertification. Science 247: 1043-1048. Tiedemann, A.R., J.O. Klemmedson and E.L. Bull. 2000. Solution of forest health problems with prescribed fire: Are forest productivity and wildlife at risk? Forest Ecology and Management 127 (1-3): 1-18. Wei, X., J.P. Kimmins and G. Zhou. 2003. Disturbances and the sustainability of long-term site productivity in lodgepole pine forests in the central interior of British Columbia: An ecosystem modeling approach. Ecological Modelling 164: 239-256. Wilson, C.A., R.J. Mitchell, L.R. Boring and J.J. Hendricks. 2002. Soil nitrogen dynamics in a fire-maintained forest ecosystem: results over a 3-year burn interval. Soil Biology and Biochemistry 34: 679-689. Wissel, C. 1984. A universal law of the characteristic return time near thresholds. Oecologia 65: 101-107. Wondzell, S.M. and J.G. King. 2003. Postfire erosional processes in the Pacific Northwest and Rocky Mountain regions. Forest Ecology and Management 178 (1-2): 75-87. Wright, R.J. and S.C. Hart. 1997. Nitrogen and phosphorus status in a ponderosa pine forest after 20 years of interval burning. Ecoscience 4(4): 526-533. |
![]()
(Back to top)
María del Carmen COBO (mccobo@ujaen.es)
is a Ph.D. student in the Department of Zoology, Botany and Ecology, University
of Jaén; she will soon defend her doctoral thesis "Small scale spatial
heterogeneity and soil-plant relationships in semiarid ecosystems from SE Spain."
She also collaborates on research in Dr. M.W. Palmer's lab in the Department
of Botany at Oklahoma State University, and is a member of the Andalusian research
group "Forest ecology and landscape dynamics" conducted by her advisor,
Dr. J.A. Carreira.
José Antonio CARREIRA (jafuente@ujaen.es) is a Professor of Ecology in the Department of Zoology, Botany and Ecology, University of Jaén. At present, he is the main researcher of the research group RNM-296 (Andalusian government catalogue) "Forest ecology and landscape dynamics," operating at the University of Jaén.
Authors' correspondence address:
Departamento de Biología Animal, Biología Vegetal y Ecología
Facultad de Ciencias Experimentales
Universidad de Jaén
23071-Jaén
SPAIN
(Back to top)
University of Jaen, RNM-296: Forest ecology and landscape dynamics
http://www.ujaen.es/investiga/rnm296/rnm296.html
This Spanish-language site provides more information on the research activities
of the co-authors and their colleagues in this research group.
About the Arid Lands Newsletter